








- Популярные
- Новые
- Акционные
- Ожидаемые
- Покупаемые
- Обсуждаемые
- Оптовый заказ
- Комплект
- Файлы
- Склады
- Позвоним

-
 Гормоны роста
Гормоны роста -
 Пептиды
Пептиды - ГОТОВЫЕ КУРСЫ ПЕПТИДОВ
- Пролонгированые пептиды
- Стимуляторы выработки гормона ро...
- Стимуляторы выработки гормона ро...
- Факторы роста (MGF/IGF)
- Стимуляторы загара
- Стимуляторы жиросжигания
- Стимуляция собственного тестосте...
- Стимуляторы нейронной активности
- Стимуляторы сексуальных функций
- Стимуляторы выносливости
- Антиэстрогены
-
 SARMs
SARMs -
 БАДы
БАДы
ДЕЙСТВИЕ ДЕЛЬТА-СОН ИНДУЦИРУЮЩЕГО ПЕПТИДА (DSIP) ПРИ ЭПИЛЕПСИИ
ДЕЙСТВИЕ ДЕЛЬТА-СОН ИНДУЦИРУЮЩЕГО ПЕПТИДА НА МЕТАБОЛИЗМ МОНОАМИНОВ ПРИ ЭКСПЕРИМЕНТАЛЬНОЙ ЭПИЛЕПСИИ
Е.Л. Доведова, Н. Д. Ещенко
Эпилепсия и эпилептоподобные судорожные состояния относятся к числу распространенных тяжелых заболеваний нервной системы; по данным ВОЗ в мире эпилепсией страдает около 40 млн человек [1]. К настоящему времени усилиями нейрохимиков, нейрофизиологов, фармакологов, а также специалистов в области молекулярной биологии и генетики накоплены сведения, позволяющие понять основные причины возникновения и механизмы развития этого заболевания и обосновать пути его лечения.
Однако клиницистам известно, что у 20-30% лиц, страдающих эпилепсией, обнаруживается нечувствительность, резистентность как к классическим давно применяемым противоэпилептическим средствам, так и к новейшим препаратам, разработанным на основе фундаментальных исследований последних лет [2]. Более того, по мере прогрессирования заболевания у больных нарастают органические расстройства, снижается чувствительность к антиконвульсантам и нейролептикам, что приводит к ослаблению лечебного эффекта, интоксикации и развитию аллергических реакций. Остается не до конца решенной и проблема побочных эффектов противоэпилептических лекарств, поскольку эти лекарства больные вынуждены принимать в течение длительного времени, в ряде случаев — пожизненно. Поэтому весьма актуальным и важным является поиск новых препаратов, применение которых позволило бы использовать более низкие дозы антиконвульсантов без ослабления их терапевтического эффекта.
Большое внимание исследователей в этом отношении привлекают различные регуляторные пептиды как эндогенные соединения, активно участвующие в регуляции многих физиологических функций. Так, установлено, что нейропептид тиролиберин в ультранизких концентрациях (10-8-10-12 моль/л) тормозит судорожные припадки у животных в разных моделях эпилепсии [3]. Положительные изменения в ЭЭГ-показателях, нормализация мозгового кровотока и снижение тяжести судорожных припадков зарегистрированы у больных при интраназальном введении тиролиберина [4].
Еще одним пептидом, который давно привлекает внимание исследователей, является дельта-сон индуцирующий пептид (ДСИП). Экспериментальное повышение его уровня в организме сопровождается изменением целого ряда физиологических функций. Открытый еще в конце 70-х годов XX в., этот пептид до сих пор во многом остается загадкой для нейрохимиков. Во-первых, в отличие от многих регуляторных пептидов для ДСИП пока не найдено белка-предшественника. Во-вторых, индукция у бодрствующих животных дельта-волновой активности мозга, характерной для ортодоксальной фазы сна (что обусловило название этого пептида), как выяснилось позднее, не является основным биологическим эффектом ДСИП. Поэтому сейчас его рассматривают как природный адаптоген широкого спектра действия [5].
К настоящему времени накоплены многочисленные доказательства тому, что ДСИП повышает устойчивость организма к различным видам стрессорных воздействий, например, к гипоксии и гипероксии, эмоциональному стрессу, гипокинезии [6, 7, 8]. Этот пептид оказывает регулирующий эффект при алкогольной мотивации, при дисфункции ней-ромедиаторных систем, в том числе катехоламинергической и холинергической [9, 10, 11].
Целью данной работы было изучение эффектов дельта-сон индуцирующего пептида на активность основных ферментов метаболизма моноаминов, выполняющих нейроме-диаторные функции, в условиях экспериментальной эпилепсии. Известно, что в развитии судорожного припадка при эпилепсии или при других формах судорожной активности большую роль играет нарушение работы моноаминергических систем — в первую очередь серотонинергической и дофаминергической [12, 13]. В связи с этим нам представлялось необходимым исследовать изменения активности основного фермента метаболизма моноаминов — моноаминоксидазы (формы МАОА и МАОБ) при выбранных экспериментальных воздействиях.
Материалы и методы исследования
Исследование выполнено на беспородных кошках массой 3,0-3,5 кг, содержавшихся в стандартных условиях вивария при свободном доступе к пище и воде. Для проведения электроэнцефалографического контроля всем кошкам заранее были вживлены электроды в различные области мозга (слуховую, зрительную, двигательную зоны коры, хвостатое ядро, срединный центр таламуса, гиппокамп). Вживление электродов проводили по методике, разработанной в Институте мозга ВНЦ психического здоровья РАМН [14].
Экспериментальную форму эпилепсии вызывали внутримышечным введением пенициллина в дозе 400 000 ед./кг массы тела (препарат калий-бензилпенициллин производства фирмы
Животные были разделены на следующие группы по 5 кошек в каждой группе: 1) контрольная, 2) животные с экспериментальной формой эпилепсии, 3) кошки, которым через 60 мин после появления выраженной эпилептиформной активности внутри-брюшинно вводили пептид дельта-сна в дозе 120 мг/кг массы тела, 4) животные, которым ДСИП в той же дозе вводили непосредственно перед инъекцией пенициллина.
Для определения биохимических показателей животных декапитировали под легким эфирным наркозом через 2 ч после начала эпилептических судорог. Методом дифференциального центрифугирования в градиенте плотности сахарозы (0,8-1,4 М) из сенсомоторной зоны коры больших полушарий и из хвостатого ядра мозга выделяли фракции «легких» и «тяжелых» синаптосом (С и D соответственно), а также фракцию свободных митохондрий нейронов. Выделение субклеточных фракций проводили, пользуясь классическим методом Е. Де Робертиса и соавторов [16].
Активность ферментов метаболизма моноаминов в изолированных субклеточных фракциях определяли спектрофотометрически. Анализ активности моноаминооксида-зы А (субстрат—серотонин) проводили по методу В. З. Горкина и соавторов [17], активность моноаминооксидазы Б (субстрат—п-нитрофенилэтиламин) определяли методом Н. Попова и соавторов [18] с некоторыми модификациями [19].
Статистическую обработку полученных экспериментальных результатов проводили с применением ^критерия Стьюдента.
Результаты исследования и их обсуждение
Данные, полученные при определении активности ферментов метаболизма моноаминов (МАОА и МАОБ) в субклеточных фракциях, изолированных из сенсомоторной коры и хвостатого ядра мозга животных четырех экспериментальных групп, приведены в табл. 1 и 2. При анализе этих результатов прежде всего хочется обратить внимание на значительное превалирование активности МАОА над активностью МАОБ во всех исследованных субклеточных фракциях. Наиболее выражено это различие во фракции «легких» синаптосом (фракция С), которая содержит основную массу синаптических окончаний катехоламинергических нейронов. Так, во фракции «легких» синаптосом, выделенных из сенсомоторной коры мозга контрольных животных, активность МАОА более чем в 12 раз превышает активность МАОБ (см. табл. 1). В такой же фракции синаптосом, изолированных из хвостатого ядра, это превышение составляет 5,1 раза (см. табл. 2).
Таблица 1. Активность моноаминоксидаз в субклеточных фракциях сенсомоторной зоны коры мозга кошек различных экспериментальных групп (M ± m; n = 5 в каждой группе)
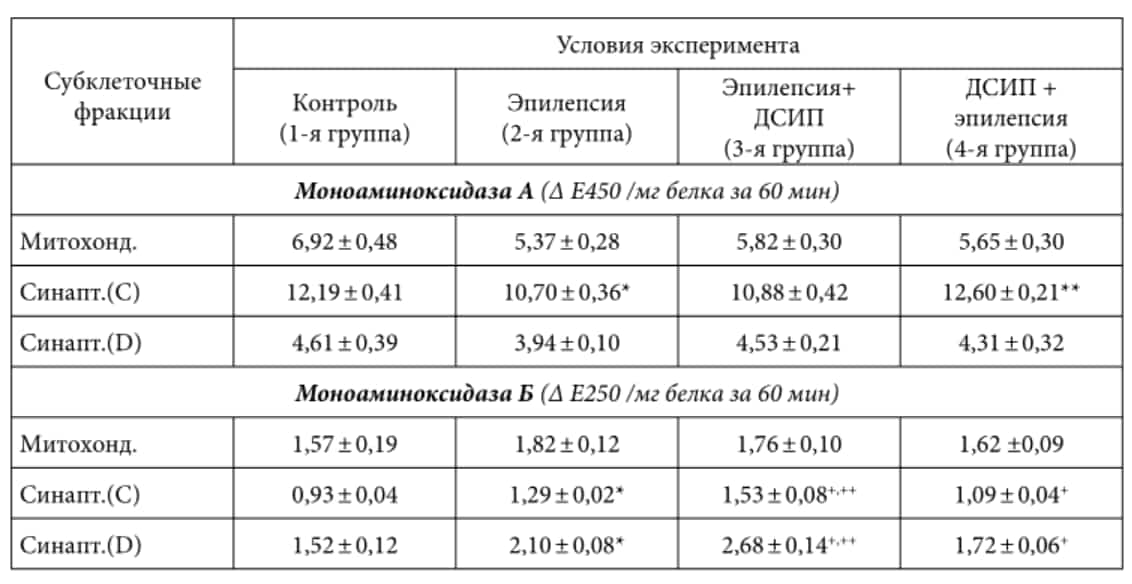
Примечание. * — различие достоверно (р < 0,05) при сравнении группы 2 (эпилепсия) с контролем (1-я группа); ** — различие достоверно (р < 0,05) при сравнении результатов, полученных во 2-й группе (эпилепсия) и 4-й группе (ДСИП + эпилепсия); + — различие достоверно (р < 0,05) при сравнении результатов, полученных во 2-й группе (эпилепсия) и 3-й группе (эпилепсия + ДСИП), а также во 2-й группе (эпилепсия) и 4-й группе (ДСИП + эпилепсия); ++ — различие достоверно (р < 0,05) при сравнении результатов, полученных в 1-й группе (контроль) и 3-й группе (эпилепсия + ДСИП).
Интересно отметить, что подобное значительное преобладание активности МАОА над активностью МАОБ описано в различных структурах мозга многих экспериментальных животных (крысы, кролики, морские свинки и др.), в то время как для ряда структур мозга человека, напротив, характерна, более высокая активность МАОБ по сравнению с МАОА. Высказано предположение, что наличие высокоактивной МАОБ (в частности, в серотонинергических нейронах) предохраняет клетку от накопления основного субстра-
та этой формы оксидазы — дофамина, который может нарушить накопление в везикулах, выброс в синаптическую щель и обратный захват серотонина [13].
Таблица 2. Активность моноаминоксидаз в субклеточных фракциях хвостатого ядра мозга кошек различных экспериментальных групп (М± m; n=5 в каждой группе)
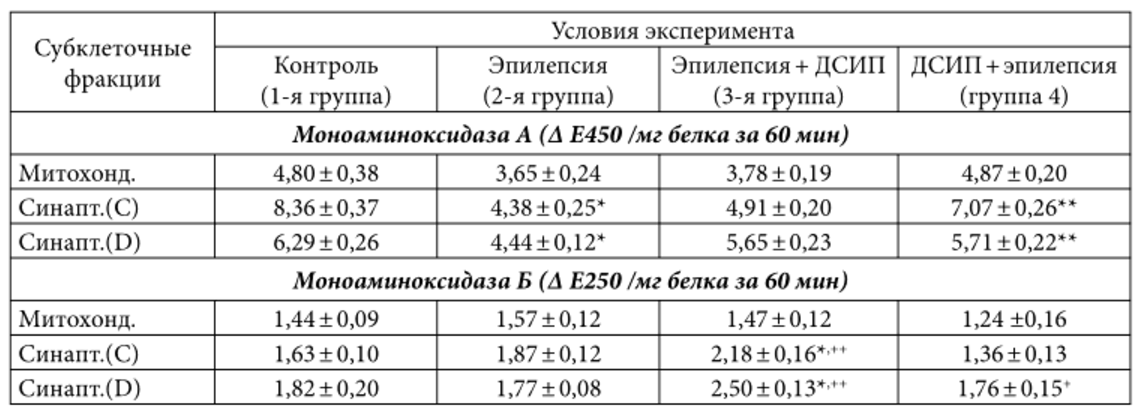
Примечание. Обозначения, как в табл. 1.
Введение животным пенициллина приводило к развитию продолжительной судорожной активности: уже через 40-45 мин на ЭЭГ появляются циклические эпилепти-формные разряды длительностью от 1 до 1,5 сек с интервалом 8-25 с. Эпилептиформная активность охватывала все структуры мозга. Такие изменения ЭЭГ регистрировались в течение 3-4 ч. На этом фоне у животных наблюдались миоклонические судороги, быстро перерастающие в эпилептический припадок.
На фоне развившейся у экспериментальных животных судорожной активности нами обнаружено снижение активности МАОА в субклеточных фракциях, выделенных как из сенсомоторной коры, так и из хвостатого ядра. Наиболее существенным было уменьшение активности фермента во фракции «легких» синаптосом из хвостатого ядра, где этот показатель падал в среднем до 48% от контрольных значений; почти на 30% снижалась активность МАОА во фракции «тяжелых» синаптосом (фракция D) из хвостатого ядра (см. табл. 2). В субклеточных фракциях из сенсомоторной коры мозга снижение активности МАОа было менее выраженным (примерно на 15-20%).
Приведенные в табл. 1 и 2 результаты указывают на то, что соотношение активности двух исследованных форм моноаминооксидазы сохраняется в структурах мозга кошек всех экспериментальных групп (животные с эпилептическими судорогами, а также кошки, которым вводили ДСИП до развития судорог или во время судорожной активности), т. е. во всех случаях активность МАОА превышала активность МАОБ
Напротив, активность другой формы моноаминооксидазы — МАОБ в исследованных структурах мозга при экспериментальной эпилепсии в субклеточных фракциях из хвостатого ядра оставалась в пределах, близких к контрольным значениям. Во фракциях синаптосом, выделенных из сенсомоторной коры, в условиях эпилептических судорог обнаружено повышение активности МАОБ на 38-40% (см. табл. 1).
Известно, что преимущественными субстратами окисления МАОА являются нор-адреналин и серотонин, в то время как МАОБ участвует главным образом в окислении дофамина. Обнаруженные нами разнонаправленные изменения активности двух форм
фермента могут указывать, во-первых, на разную степень вовлеченности серотонин-, норадреналин- и дофаминергических систем мозга в развитие эпилептического статуса; во-вторых, на нарушение взаимоотношений этих медиаторных систем в условиях судорожной активности.
Введение животным ДСИП на фоне уже развившегося эпилептического припадка (3-я экспериментальная группа) не приводило к изменениям характеристик эпилепти-формных разрядов на ЭЭГ. На субклеточном уровне наблюдалась некоторая тенденция к нормализации значений активности МАОа в сенсомоторной коре и в хвостатом ядре. В то же время активность МАОБ во фракциях синаптосом обоих структур мозга была выше не только по сравнению с контролем, но и превышала значения, найденные в мозге кошек с эпилептическим статусом (см. табл. 1 и 2).
У животных, которым инъекции ДСИП делали предварительно до введения пенициллина и, следовательно, до развития судорожной активности (4-я экспериментальная группа животных), мы наблюдали иной характер ЭЭГ. Эпилептиформные разряды распространялись не на все структуры мозга; они охватывали структуры последовательно: вначале слуховую, зрительную зоны коры и гиппокамп, а затем сенсомоторную зону коры, хвостатое ядро и срединный центр таламуса. В хвостатом ядре и срединном центре таламуса проявлялись периоды (длительностью 5-8 с) медленно-волновой активности. Миоклонические сокращения мышц у животных этой группы не наблюдались.
Предварительное введение ДСИП не только препятствовало развитию судорожного состояния у экспериментальных животных 4-й группы, но и приводило к некоторой нормализации активности обеих форм моноаминооксидазы в субклеточных фракциях исследованных структур мозга. Так, активность МАОА и МАОБ во фракциях синаптосом из сенсомоторной коры мозга была близка к контрольным значениям (см. табл. 1); статистически значимых отличий при сравнении этих двух групп нет. Во фракциях си-наптосом, выделенных из хвостатого ядра животных данной группы, активность МАОБ также была на уровне контроля. Активность МАОА в «легких» (фракция С) и «тяжелых» (фракция D) синаптосомах из хвостатого ядра, хотя и не достигала контрольных значений, но тем не менее была выше, чем в мозге животных в условиях эпилептических судорог (см. табл. 2).
Таким образом, полученные нами экспериментальные данные говорят о том, что нормализующее действие пептида дельта-сна проявляется лишь при условии введения его до развития судорожного состояния. Эти результаты согласуются с имеющимися в литературе сведениями о том, что именно превентивное введение ДСИП позволяет наиболее четко выявить его свойства адаптогена при различных стрессорных состояниях; в этих условиях данный пептид способствует замене стресс-реакции на более эффективные и экономичные адаптационные стратегии [8, 21].
Несмотря на то, что участие ДСИП в регуляции активности моноаминоксидаз обнаружено довольно давно [21], биохимические механизмы, обеспечивающие реализацию адаптогенного эффекта ДСИП, до сих пор окончательно не расшифрованы.
Экзогенный дельта-сон индуцирующий пептид легко преодолевает гематоэнцефа-лический барьер и достаточно равномерно распределяется в структурах мозга [22]. Период полураспада ДСИП, как и многих других нейропептидов, измеряется минутами [5], в то время как время его последействия может быть гораздо более длительным, судя по пролонгированному антистрессорному эффекту. Это позволяет предположить, что ан-тистрессорное действие ДСИП определяется не только его непосредственным влиянием
на конкретный фермент, но, скорее всего, целым каскадом биохимических реакций, запускаемых этим регуляторным пептидом. Такое предположение подтверждается данными об участии ДСИП в регуляции экспрессии генов раннего ответа, в частности гена c-fos [23], который, наряду с другими «ранними генами» и транскрипционными факторами (CREB, AP-1 и др.), способствует изменению программы клеточного ответа вплоть до включения защитных (противосудорожных) механизмов.
Кроме того, вполне возможно, что определенный вклад в реализацию нормализующего эффекта ДСИП при экспериментальной эпилепсии вносит регуляция им активности протеолитических ферментов. Действие пептида при этом направлено на элиминацию образующихся в ходе развития стресс-реакции белков с поврежденной структурой. Подобный аспект в действии ДСИП обнаружен А. М. Менджерицким и соавторами [24], которые показали, что предварительное введение пептида крысам вызывает повышение активности катепсина Д и калпаина типа I в синаптосомах и препятствует изменениям ультраструктуры нейронов, вызываемым гипокинезией.
Полученные нами результаты позволяют сделать заключение о том, что эпилептические судороги, вызванные введением пенициллина экспериментальным животным, сопровождаются снижением моноаминоксидазной активности (форма МАОА), наиболее выраженным во фракциях синаптосом хвостатого ядра, в то время как активность МАОБ оставалась близкой к контрольным значениям (в хвостатом ядре) или несколько возрастала (в сенсомоторной коре мозга). Это указывает на нарушение функционирования моноаминергических медиаторных систем мозга. Дельта-сон индуцирующий пептид способен предотвратить изменения активности моноаминоксидаз только в случае, когда его вводили животным превентивно, до развития эпилептических судорог.
Литература
1. Gibbs J. W., McNamara J. O. The epilepsies. Phenotype and mechanisms // Basic Neurochemistry / ed. by G. J. Siegel. Elsevier Acad.Press, 2006. L. P. 629-637.
2. Вайнтруб М. Я. Эпилепсия как хронические дизритмии головного мозга. М.: Эслан, 2000. 200 с.
3. Чепурнов С. А., Аббасова К. Р., Аристов А. В., Бердиев Р. К. Тиролиберин в регуляции фокальной эпилепсии // Нейрохимия. 2008. Т. 25, № 1-2. С. 70-81.
4. Chepurnov S. A., Chepurnova N. E., Asanova L. M., Ashmarin I. P. Neuropeptide thyroliberin may prevent epileptogenic activity in childrens // Epileptologia (Warsawa). 1999. Vol. 7, Suppl. N 1. P. 86-87.
5. Ковальзон В. М., Стрекалова Т. В. Дельта-сон индуцирующий пептид — тридцать лет спустя // Нейрохимия. 2006. Т. 23, № 4. С. 276-282.
6. Альперович Д. В., Лысенко А. В., Мационис А. Э., Менджерицкий А. М. Некоторые биохимические механизмы нейропротекторного эффекта ДСИП при гипоксии // Hypoxia Medical J. 1997. N 4. С. 3-7.
7. Лысенко А. В., Альперович Д. В., Ускова Н. И., Михалева И. И. и др. Сравнительное изучение эффективности применения ДСИП для коррекции функционально-метаболических сдвигов в условиях гипоксии и физической нагрузки // Нейрохимия. 1999. Т. 16, № 1. С. 37-44.
8. Лысенко А. В., Ускова Н. И., Альперович Д. В., Менджерицкий А. М. Особенности метаболического обеспечения адаптогенных эффектов ДСИП и пирацетама при гипероксии // Биохимия.
1999. Т. 64, № 6. С. 62-69.
9. Войтенко Н. Н. Влияние дельта-сон индуцирующего пептида и эмоционального стресса на агрессивность в тесте резидент-интрудер и активность моноаминоксидазы в мозге мышей С57BL // Нейрохимия. 2006. Т. 23, № 3. С. 199-207.
10. Войтенко Н. Н. Влияние дельта-сон индуцирующего пептида и эмоционального стресса, примененных пренатально, на моноаминоксидазу мозга и на поведение взрослых потомков-сам-цов мышей DD // Нейрохимия. 2008. Т. 25, № 3. С. 227-232.
11. Хрусталев Д. А., Доведова Е. Л., Ещенко Н. Д. Эффекты ДСИП на фоне дисфункции моно-аминергических систем в структурах мозга крыс линий Вистар и Август // Вестн. С.-Петерб. ун-та. 2007. Сер. 3, вып. 1. С. 45-49.
12. Шоу Д. А. Нейротрансмиттерные системы. М.: Медицина. 1982. 211 с.
13. Shih J. C., Hensler J. G. Serothonin // Basic Neurochemistry / ed. by G. J. Siegel. Elsevier Acad.Press. L., 2006. P. 227-248.
14. Попова Н. С., Адрианов О. С., Весков Р., Ракич Л. Влияние пептида дельта-сна на межцен-тральную интеграцию в условиях экспериментальной эпилепсии // Бюл. экспер. биол. мед. 1989. Т. 108, № 8. С. 147-150.
15. Rakich L., Rusich A., Veskov R. Morpho-functional analysis of epileptogenic activity in cat brain induced by penicillium injection // Adv. Biosci. 1988. Vol. 70. P. 133-136.
16. De Robertis E., Peligrino D. A., Rodriges O.D., Salganicoff L. Cholinergic and non-cholinergic nerve endings in rat brain // J. Neurochem. 1962. Vol. 9, N 1. P. 23-35.
17. Горкин В. З., Веревкина И. В., Гриднева Л. И., Петров И. А. Методы исследования активности и специфического торможения моноаминоксидаз митохондрий // Современные методы в биохимии. М., 1968. Т. 2. С. 155-157.
18. Popov N., Rosier V., Thiemann C., Mathies H. Eine empfindiche Methode zur Bestimmung der Monoaminnnnoxydase Activitat im Gewebe durch Aldehydsemikarbazon-Messung // Acta Biol. Med. Germ. 1971. Bd 26. S. 239-241.
19. Доведова Е. Л., Хрусталев Д. А. Сравнительная характеристика ферментных систем обмена нейромедиаторов в мозге крыс Вистар и Август при различных сроках воздействия амфетамина in vivo // Нейрохимия. 2007. Т. 24, № 2. С. 150-155.
20. Белякова Е. И., Колмакова Т. С. Влияние пептида дельта-сна на моноаминергические системы мозга крыс в начальный период ноцицептивного воздействия // Нейрохимия. 2007. Т. 24, № 2. С. 156-160.
21. Доведова Е.Л., Ашмарин И. П. Действие пептида дельта-сон на активность моноаминокси-даз и ацетилхолинэстеразы в субклеточных фракциях из различных образований мозга крыс // Бюл. эксперим. биол. мед. 1982. Т. 93, № 5. С. 56-58.
22. Zlokovic B. V., Segal M. V., Davson H. Autoradiographic investigations of DSIP distribution in rat brain // Peptides. 1988. Vol. 9. P. 533-538.
23. Умрюхин П. Е. Циклогексемид блокирует подавление пептидом, вызывающим дельта-сон, экспрессии раннего гена c-fos в паравентрикулярных ядрах гипоталамуса // Бюл. экспер. биол. мед.
2000. Т. 134, № 9. С. 254-256.
24. Менджерицкий А. М., Лысенко А. В., Ускова Н. И., Мационис А. Э. Роль протеолитических ферментов в механизме действия дельта-сон индуцирующего пептида в норме и в условиях гипокинезии // Нейрохимия. 1996. Т. 13, № 1. С. 23-31.
Статья поступила в редакцию 20 декабря 2010 г.
Научная библиотека КиберЛенинка: https://cyberleninka.ru/article/n/deystvie-delta-son-indutsiruyuschego-peptida-na-metabolizm-monoaminov-pri-eksperimentalnoy-epilepsii#ixzz4O6rg9uwF
